|
Eco-epidemiología del virus
encefalitis St. Louis en Córdoba, Argentina.
Lorena I. Spinsanti, Luis A. Diaz, Marta S. Contigiani
Revista
Facultad de Ciencias Médicas 2009; 66(Supl.1):52-59
Laboratorio de Arbovirus,
Instituto de Virología “Dr. J. M. Vanella”, Facultad de
Ciencias Médicas, Universidad Nacional de Córdoba.
Introducción
El virus Encefalitis de St. Louis (VESL), pertenece al
género Flavivirus, familia Flaviviridae y en base a sus
determinantes antigénicos forma parte del serogrupo
Encefalitis Japonesa junto con otros Flavivirus de
importancia médica y veterinaria como West Nile, Encefalitis
Japonesa, Murray Valley (1).
El VESL se aisló por primera vez en el año 1933 a partir del
cerebro de un paciente durante un brote de encefalitis en la
ciudad de Saint Louis, EE.UU. Este virus se halla
ampliamente distribuido en América, reportándose actividad
desde Canadá hasta el sur de Argentina. En los EE.UU. ha
sido una de las principales causa de epidemias de
encefalitis por arbovirus hasta la introducción del virus
West Nile (VWN) en el año 1999 (2). Sin embargo, en el resto
del continente americano los casos clínicos observados por
este virus han sido principalmente de enfermedad febril, sin
registro de brotes (3).
Las manifestaciones clínicas generadas por la infección del
VESL se han agrupado en tres síndromes principales:
encefalitis, meningitis aséptica y cefalea febril. La
mayoría de las infecciones con el VESL en personas son
asintomáticas y la susceptibilidad de adquirir el síndrome
encefálico aumenta con la edad del paciente, reportándose un
70% de casos en personas mayores de 75 años durante las
epidemias en EE.UU. (4)
El diagnóstico presuntivo se basa en la detección de
anticuerpos IgM por MAC-ELISA; estos anticuerpos pueden
persistir hasta un año. El diagnóstico definitivo es la
demostración del aumento o la disminución de anticuerpos
entre el suero agudo y convaleciente (seroconversión) por la
técnica de Neutralización (NT). (4)
Ciclos de Transmisión y Ecología
El VESL es mantenido y amplificado en los EE.UU. por
transmisión horizontal en un ciclo enzoótico primario que
involucra aves silvestres (principalmente paseriformes –gorriones
Passer domesticus- y Columbiformes –palomas Zenaida macroura)
y mosquitos del género Culex (4). El hombre es un hospedador
accidental. En la zona este de los EE.UU., Culex pipiens y
Culex quinquefasciatus son los mosquitos vectores primarios;
en estas áreas el virus circula en forma urbana y suburbana.
En contraste, en la zona oeste, el virus circula en áreas
rurales, principalmente irrigadas donde se reproduce el
mosquito vector Culex tarsalis. (4)
Los ciclos de transmisión que mantienen activo al VESL en
América Central y del Sur están poco caracterizados. En esta
región el virus ha sido aislado a partir de 11 géneros
diferentes de mosquitos, incluyendo Cx. nigripalpus y Cx.
quinquefasciatus (3). Existen fuertes evidencias a favor de
las aves como hospedadores del VESL. Varias cepas virales
han sido aisladas de 27 especies de aves, incluyendo
cormoranes, garzas, palomas, zorzales y celestinos. También
se han encontrado mamíferos silvestres (microroedores,
perezosos) y domésticos infectados, aunque las evidencias a
favor de su rol como hospedadores son escasas (3).
Las distintas cepas aisladas del VESL poseen variabilidad
biológica y genética a lo largo de su rango de distribución.
Monath y col. (5) detectaron la existencia de cepas
virulentas y atenuadas, mientras que estudios genéticos
identificaron la presencia de diversos genotipos con
distribución geográfica heterogénea (6,7). En base a la
secuenciación completa del gen de la envoltura, Kramer y
Chandler (8) clasificaron a las cepas del VESL en 7
genotipos (I, II, III, IV, V, VI y VII), confirmando la
variabilidad genética y biológica detectada años anteriores.
Antecedentes en Argentina
Aislamientos virales:
El VESL es endémico en Argentina y posee una amplia
distribución geográfica, detectándose actividad tanto en
regiones subtropicales (Misiones, Chaco, Formosa, Tucumán)
como templadas (Córdoba, Corrientes, Entre Ríos, Mendoza,
Santa Fe y Santiago del Estero) del país (9).
En Argentina se han aislado un total de 12 cepas desde el
año 1963 hasta el 2005. Las fuentes de aislamiento son
variadas y abarcan mosquitos (8 cepas), roedores (2 cepas) y
humanos (2 cepas), colectadas de diferentes provincias.
(9,10,11,12) Los detalles correspondientes a todas las cepas
se encuentran en la Tabla I.
Estudios en vectores:
Durante los años 2001-2004 se realizó en la ciudad de
Córdoba un estudio con el objetivo de detectar la
variabilidad genética natural circulante del VESL y analizar
los patrones de actividad viral. Se detectaron un total de
20 pools de mosquitos infectados con VESL en 8 especies
diferentes de mosquitos, para la mayoría de las cuales (Ae.
aegypti, Ae. scapularis, An. albitarsis, Cx. apicinus) no
existían antecedentes de infección por este virus en
Argentina ni en América del Sur (13). Las 4 especies más
abundantes y frecuentemente infectadas fueron Cx. interfor,
Cx. quinquefasciatus, Ae. albifasciatus y Ae. scapularis. Se
detectó una marcada estacionalidad en la actividad temporal
del VESL, coincidente con los patrones temporales de
actividad en regiones templadas de América del Norte (4). La
actividad enzoótica del VESL para la ciudad de Córdoba se
extendería desde Noviembre hasta Abril (verano-otoño), con
picos anuales en Febrero-Marzo y ocasionalmente en Diciembre,
dependiendo de las condiciones climáticas que influyen sobre
las poblaciones de mosquitos vectores. Los picos máximos de
actividad viral se registraron en el período de noviembre de
2002 a marzo de 2003, coincidiendo por lo general con la
predominancia de las comunidades de mosquitos de Culex
interfor y Culex quinquefasciatus (13). Se detectaron cepas
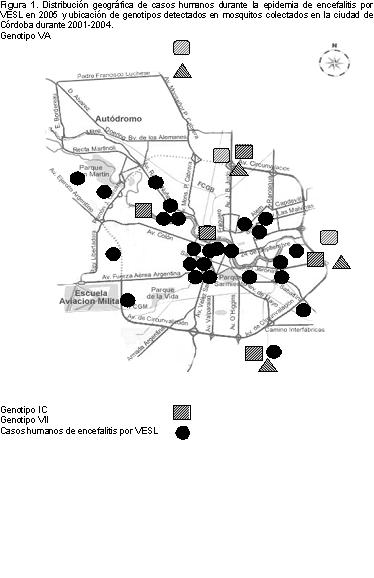
pertenecientes a 3 genotipos: IC, VA y VII, siendo el
segundo dominante sobre los restantes minoritarios (14). La
existencia de un genotipo dominante podría deberse a cepas
del virus que son más viremogénicas, generando viremias
elevadas en aves e infectando un mayor número de mosquitos,
permitiendo una mayor circulación y dispersión de las cepas.
Durante la ocurrencia del brote por VESL en Córdoba, Díaz y
col. (12) aislaron dos cepas del VESL pertenecientes al
genotipo III a partir de mosquitos Cx. quinquefasciatus
coleccionados en el domicilio de uno de los pacientes con
encefalitis. Este genotipo emergió en Córdoba en el año
2005, después de su última detección en la provincia de
Santa Fe, 26 años atrás (9).
Mitchell et al. (15) determinaron la transmisión vectorial
de las cepas autóctonas, 78V-6507 y 79V-2533, del VESL
aisladas en la provincia de Santa Fe por mosquitos Cx.
quinquefasciatus colectados en la misma provincia, poniendo
en evidencia el probable rol de estos mosquitos como
vectores del VESL en nuestro país. Recientemente se detectó
que poblaciones locales de mosquitos Culex quinquefasciatus
son altamente susceptibles de infectarse por vía oral con la
cepa epidémica CbaAr-4005 (73,10% de infección) y son
capaces de transmitir vectorialmente el virus entre pollitos
Gallus gallus (13).
En síntesis, los estudios de competencia vectorial,
abundancia poblacional de mosquitos y aislamiento viral
confirman que Culex quinquefasciatus es el principal
mosquito vector del VESL en Córdoba. Otras especies como
Culex interfor, Ae. albifasciatus y Ae. scapularis deberían
ser analizadas como vectores alternativos del VESL.
Estudios en hospedadores:
Existen antecedentes de actividad enzoótica del VESL en
diversas especies de aves silvestres y domésticas en
provincias del centro y norte de Argentina. En la provincia
de Córdoba, Sabattini (11) detectó anticuerpos inhibidores
de la hemoaglutinación en palomas Torcazas (Zenaida
auriculata) entre 1964-1965 en montes cercanos a la
localidad de Piquillín (este de la provincia de Córdoba).
Doce años más tarde, Monath et al. (16) detectaron una
prevalencia de anticuerpos neutralizantes (AcNT) del 13,6%
en la misma especie y en el mismo lugar, dando cuenta de la
endemicidad del VESL en la región. Individuos de otra
especie de paloma, Torcacita (Columbina picui), también se
encuentran frecuentemente infectados en regiones templadas
del país. Entre 1977 - 1980, se detectó la presencia de AcNT
en especies de aves pertenecientes a las familias:
Furnaridae (4,6%; 11/237), Columbidae (5,9%; 14/236),
Tyranidae (2,4%; 4/170), Fringilidae (4,9%; 8/164),
Icteridae (1,6%; 1/64), Ardeide (4,7%; 2/43) y Phytotomidae
(7,1%; 1/14) colectadas en las provincias de Chaco, Córdoba,
Corrientes y Santa Fe (16). A diferencia de lo que ocurre en
EE.UU., ninguno de los 230 sueros de gorriones analizados en
este estudio presentó AcNT, marcando una diferencia
ecológica importante entre ambos países.
En Córdoba, las aves de hábitos urbanos y periurbanos como
palomas Torcazas, palomas Torcacitas, Horneros (Furnarius
rufus), Cacholotes (Pseudoseisura lophotes), Benteveos (Pitangus
sulphuratus) y Gorriones (Passer domesticus) se detectaron
infectados en períodos epidémicos, confirmando el rol de las
aves en el mantenimiento y amplificación del VESL en nuestro
país (17). El 99% de las aves colectadas en la ciudad de
Córdoba el año previo a la epidemia no tenían AcNT contra el
VESL (18). Esta baja actividad viral observada en aves
coincidió con la escasa, casi nula, actividad del virus
detectada en mosquitos para ese mismo período (Enero-Abril
2004). Tampoco se detectó infección por VESL en gallinas
centinelas. En el período siguiente (Enero-Abril 2005), la
prevalencia de AcNT para el VESL en aves registró un aumento
de 11 veces (8%), indicando activa circulación del virus en
aves silvestres en cuatro sitios de muestreo de la ciudad de
Córdoba, correlacionándose con el registro de casos de
encefalitis durante la epidemia. (13) La ausencia de
actividad viral en aves silvestres y gallinas centinelas,
previo a una epidemia por VESL, fue registrada en los
departamentos de Los Angeles, Orange y Riverside
(California, EE.UU.) en 1984 (19), características similares
a las evidencias aquí presentadas y registradas en la ciudad
de Córdoba, previo a la epidemia de 2005.
Los datos de viremia e índices de competencia de hospedador
obtenidos por Diaz (13,20), indican que las 2 especies de
palomas (Torcacita y Torcaza) (orden Columbiformes) son los
principales hospedadores aviares. A modo comparativo, las
Torcazas y Torcacitas generan 15 y 3 veces, respectivamente,
más mosquitos infecciosos que un gorrión. El rol de los
gorriones es poco importante, teniendo una actuación casi
despreciable en el mantenimiento del virus, al igual que los
tordos músico (Agelaioides badius), tordo renegridos (Molothrus
bonariensis) y paloma ala manchada (Patagioenas maculosa)
(13). Indicando diferencias ecológicas respecto a los ciclos
de mantenimiento y transmisión entre Argentina y EE.UU.
Epidemiología
Actividad epidémica del VESL en población humana:
Desde Enero del año 2002 a Diciembre de 2007 se buscó la
detección del VESL o sus anticuerpos en 728 pacientes con
síndromes febriles y/o neurológicos provenientes de la
provincia de Córdoba. Se detectaron anticuerpos IgM,
indicadores de infección reciente por VESL, en 68 pacientes.
Los meses de mayor ocurrencia de la enfermedad fueron desde
Enero a Abril. La mayoría de los pacientes afectados fueron
de sexo masculino, edad promedio 44 años y procedentes de la
Capital de la provincia de Córdoba (21). El síndrome más
observado fue la encefalitis. De acuerdo al criterio de
confirmación, 19 casos fueron confirmados como infección por
VESL.
El primer caso se detectó en Febrero del año 2002 en la zona
central de la ciudad de Córdoba (Barrio San Martín) (22),
coincidente con un pico de abundancia de mosquitos Culex
quinquefasciatus (23). Luego de dos años donde no se
detectaron casos, en el verano del año 2005, ocurre en
Córdoba el primer brote de infección por VESL en Sudamérica
y en Argentina. La curva epidémica de los 47 casos mostró
que el brote comenzó en enero del año 2005 con un pico en
febrero y un pico mayor en la semana del 6 al 12 de marzo,
finalizando en mayo (24). Del total de casos notificados, 39
ocurrieron en la capital de la provincia de Córdoba, 3 en el
departamento Colón (Río Ceballos, Villa Allende y Colonia
Tirolesa), 4 casos en el departamento Río Segundo (Villa del
Rosario, Pilar) y uno en el departamento San Javier (Villa
Dolores). En la capital de Córdoba, la distribución
geográfica de los casos fue uniforme. Entre los barrios
donde se registraron casos, el barrio Villa Belgrano, tuvo
la tasa de ataque más alta donde cada 10.000 hab 5 tuvieron
riesgo de enfermarse. Otros barrios con tasas elevadas
fueron Bella Vista, Maipú 2° sección, Renacimiento, Villa
Adela y Parque Jorge Newbery (rango 3-4/10.000 hab) (24).
En cuanto a la clínica de los pacientes, los principales
signos y síntomas que involucraron al Sistema Nervioso
Central incluyeron cefalea, depresión del sensorio,
desorientación temporo-espacial, temblores y cambios en el
nivel de conciencia. Se encontró una asociación
significativa entre la edad y la severidad de la enfermedad
con un coeficiente de Spearman igual a 0,74. Los hallazgos
del líquido cefalorraquídeo fueron típicos de una infección
viral (21)
De los 47 casos de infección por VESL, 45 fueron
hospitalizados y 2 fueron tratados en forma ambulatoria. La
edad media de los pacientes fue 47,8 ± 24,6 años (rango
7-87años). La mayoría de los casos tuvieron entre 10 y 60
años (52%), grupo etario que comprende el 70% de la
población total; 19 pacientes (40%) eran mayores de 60 años.
La mayor tasa de ataque fue observada en las personas
mayores de 60 años. El 64% de los pacientes fueron de sexo
masculino. El 80% de los casos de encefalitis y
meningoencefalitis fue en personas menores de 20 años y el
95% en mayores de 60 años. La tasa de mortalidad fue 19% (uno
fue de 25 años y 8 fueron mayores de 50 años). El 40% de los
pacientes eran jubilados y/o amas de casa (24).
Factores de riesgo e información ambiental:
La prevalencia de AcNT de los individuos que asistieron a
los distintos Centros de Atención Primaria de la Salud
(CAPS) de la ciudad de Córdoba, antes del brote, fue similar
a la previamente encontrada en los años 1998-2000 (13.3% en
el año 2002 y 12.9% en el año 2004). Los individuos que se
atendieron en el CAPS Nº58 (Bº Maldonado) demostraron la
prevalencia de AcNT más alta. En esta población el riesgo de
infección fue tres veces mayor que en la del CAPS Nº40 (Bº
Las Flores). Estos resultados coincidieron con los hallazgos
de Díaz y col. (13, 17, 23) que describen una mayor tasa de
infección en aves y una mayor abundancia de mosquitos en
esta zona, debido probablemente a la presencia de lagunas y
depósitos de basura, que proveen sitios de cría para C.
pipiens quinquefasciatus. Por esta razón, los individuos con
viviendas cercanas a basurales tuvieron 2,5 veces más chance
de infección. La práctica de actividades externas en la
noche incrementó la posibilidad de infección debido
probablemente a los hábitos nocturnos del mosquito Culex. En
el año 2005, durante la ocurrencia del brote, la prevalencia
total de AcNT detectada en tres CAPS fue la más alta de los
últimos años (22,1%). El riesgo de infección fue el doble en
el año 2005 que en el 2004. Nuevamente el CAPS Nº58 presentó
la mayor prevalencia (31%). Futuros estudios que evalúen
sistemas de drenaje, nivel educativo, conocimiento sobre la
presencia del mosquito, etc, serán necesarios para
incrementar el entendimiento de los efectos de la
interacción humano-ambiente que hacen que estos individuos
tengan un mayor riesgo de infección con el VESL que otras
poblaciones de la ciudad donde la prevalencia es menor (rango
8%-13%). (25)
Por otra parte, mediante el uso de información espacial y
datos epidemiológicos derivados del brote, se construyeron
modelos predictivos y mapas de riesgo de infección para la
ciudad de Córdoba. Se encontró una asociación directa entre
el número de humanos infectados y la cercanía a fuentes de
vegetación vigorosa y densa. El mapa de susceptibilidad
ambiental para la incidencia de infección con VESL clasificó
el 66% de la ciudad bajo riesgo medio, medio alto y alto y
34% con riesgo bajo o sin riesgo de infección. Del total de
los casos ocurridos durante la epidemia de 2005, el 94% de
los mismos fueron notificados en áreas predichas por el
modelo como de riesgo medio, medio alto y alto. (26)
Conclusión
Aún se desconocen las causas por las cuales el VESL re
emergió en el centro de Argentina. Nuestros resultados
apoyan la hipótesis que la introducción de un nuevo genotipo
(genotipo III) con un mayor potencial viremogénico en una
nueva zona (ciudad de Córdoba) podría ser una de las causas
de la re-emergencia.

Bibliografía
1. ICTVdB Management. 00.026.0.01. Flavivirus. In: ICTVdB -
The Universal Virus Database, version 4. Büchen-Osmond, C.
(Ed), Columbia University, New York, USA.2006.
2. Jia, X Y, Briese, T, Jordan, I, et al. Genetic analysis
of West Nile New York 1999 encephalitis virus. Lancet;1999,
354: 1971-1972.[Abstract]
3. Spence, L, P. St. Louis encephalitis in tropical America.
In: St.Louis encephalitis. Monath TP. Ed. Washington, DC:
APHA,1980,p 451-471.
4. Reisen, W K. Epidemiology of St. Louis encephalitis
virus. Adv Virus Research; 2003, 61: 139-183.
5. Monath, T P, Cropp, C B, Bowen, G S, Kemp, G E, Mitchell,
C J, Gardner J J. Variation in virulence for mice and rhesus
monkeys among St. Louis encephalitis virus strains of
different orgin. Am J Trop Med Hyg; 1980, 29: 948-962.[Abstract]
6. Trent, D W, Monath, T P, Bowen, G S, Vorndam, A V, Cropp,
B C, Kemp G E. Variation among strains of St. Louis
encephalitis virus: Basis for a genetic, pathogenetic and
epidemiological classification. Annals NY Acad Sci ;1980,
354: 219-237.
7. Trent, D W, Grant, J A, Vorndam, A V, Monath T P. Genetic
heterogeneity among Saint Louis encephalitis virus isolates
of different geographic origin. Virology; 1981, 114:
319-332.
8. Kramer, L D, Chandler L J. Phylogenetic analysis of the
envelope gene of St. Louis encephalitis virus. Arch Virol;
2001, 146: 2341-2355.[Abstract]
9. Sabattini, M S, Avilés, G, Monath T P. Historical,
epidemiological and ecological aspects of arbovirus in
Argentina: Flaviviridae, Bunyaviridae and Rhabdoviridae. In
“An Overview of Arbovirology in Brazil and neighboring
countries”. Travassos da Rosa APA, Vasconcelos PFC,
Travassos da Rosa JFS, Eds. Belem, Brazil: Instituto Evandro
Chagas, 1998, p.:113-134.
10. Mitchell, C J, Monath, T P, Sabattini, M S, Cropp, C B,
Daffner, J F, Calisher, C H, Jakob, W L, Christensen H A.
Arbovirus investigations in Argentina, 1977-1980. II.
Arthropod collections and virus isolations from argentine
mosquitoes. Am J Trop Med Hyg; 1985, 34: 945-955.[Abstract]
11. Sabattini MS. Arbovirus del grupo B en la República
Argentina. Su estudio en aves y roedores. Tesis doctoral.
Facultad de Ciencias Químicas. Universidad Nacional de
Rosario, Argentina, 1969, p:.96.
12. Diaz L A, Ré V, Almirón W R, Farías A, Vázquez A,
Sanchez-Seco M P, Aguilar J, Spinsanti, L Konigheim, B
Visintin, A, García J, Morales M A, Tenorio A, Contigiani M.
Genotype III Saint Louis Encephalitis virus outbreak,
Argentina, 2005. Emerg Infect Dis; 2006, 12: 1752-1754.[Full
Text]
13. Díaz, LA. Patrones de actividad y estacionalidad del
virus St. Louis Encephalitis (Flavivirus, Flaviviridae) en
la ciudad de Córdoba. Tesis Ciencias Biológicas. Facultad
Ciencias Exactas, Físicas y Naturales, 2009, p: 194.
14. Diaz, L A, Farias, A A, Ré, V, Vazquez, A, Tenorio, A,
Almirón, W R, Contigiani, M S. Distribución espacio-temporal
de genotipos del virus St. Louis encephalitis (Flavivirus,
Flaviviridae) en la ciudad de Córdoba entre 2001-2003. XXVII
Reunión Científica Anual de SAV. Diciembre 2007. Vaquerías,
Córdoba, Argentina.
15. Mitchell, C J, Monath, T P, Sabattini, M S. Transmission
of St. Louis encephalitis virus from Argentina by mosquitoes
of the Culex pipiens (Diptera: Culicidae) complex. J Med
Entomol; 1980, 17: 282-287.
16. Monath, T P, Sabattini, M S, Pauli, R, Daffner, J F,
Mitchell, C J, Bowen, G S, Cropp, C B. Arbovirus
investigations in Argentina, 1977-1980. IV Serological
surveys and sentinel equine program. Am J Trop Med Hyg;
1985, 34: 966-975.[Abstract]
17. Díaz, L A, Konigheim, B S, Aguilar, J J, Spinsanti, L I,
Castillo, C, Ortiz Bergia, S, Zarco, A, Villafañe, N,
Almiron, W R, Contigiani, M S. Actividad del virus
Encefalitis San Luis (Flavivirus) en aves durante una
epidemia de encefalitis en Córdoba, Argentina, 2005. Acta
Bioquim Clin Latinoam; 2006, 3: 298.
18. Diaz, L A, Spinsanti, L I, Aguilar, J, Occelli, M,
Boris, A S, Reynoso, R, Kambic A, Obredor, C, Almirón, W R,
Contigiani, M S. Circulación enzoótica del virus Encefalitis
San Luis (ESL) (Flavivirus) en comunidades de aves en la
ciudad de Córdoba en el año 2004. Rev Argent Microb;2005,
37: 33.
19. Murray, R A, Hable, L A, Mackey, K J, Wallace, H G,
Peck, B A, Mora, S J, Ginsberg, M M, Emmons, R W.
Epidemiologic aspects of the 1984 St. Louis encephalitis
epidemic in southern California. Proc Calif Mosq Vect Contr
Assoc; 1985, 53: 5-9.
20. Diaz, L A, Occelli, M, Ludueña Almeida, F, Almirón, W R,
Contigiani, M S. Eared Dove (Zenaida auriculata, Columbidae)
as host for St. Louis encephalitis virus (Flaviviridae,
Flavivirus). Vector Borne Zoonotic Dis; 2008, 8: 277-282.[Abstract]
21. Spinsanti, L I. Epidemiología del virus Encefalitis de
St. Louis en la provincial de Córdoba. Tesis doctoral.
Facultad Ciencias Médicas. Universidad Nacional de Córdoba.
2009
22. Spinsanti L, Basquiera A L, Bulacio S, Somale V, Kim S
C, Ré V, Rabbat D, Zárate A, Zlocowski J C, Mayor C Q,
Contigiani M, Palacio S. St. Louis encephalitis in
Argentina: the first case reported in the last seventeen
years. Emerg Infect Dis; 2003, 9: 271-273.
[Full Text]
23. Diaz, L A, Almiron, W R, Ludueña Almeida, F, Spinsanti,
L I, Contigiani M S. Vigilancia del virus Encefalitis de San
Luis y mosquitos (Diptera: Culicidae) en la Provincia de
Córdoba, Argentina. Entomol Vectores ;2003, 10:551-566.
24. Spinsanti, L I, Díaz, L A, Glatstein, N, Arselán, S,
Morales, M A, Farías, A A, Fabbri, C, Aguilar, J J, Ré, V,
Frías, M, Almirón, W R, Hunsperger, E, Siirin, M, Da Rosa, A
T, Tesh, R B, Enría, D, Contigiani, M S. Human outbreak of
St. Louis encephalitis detected in Argentina, 2005. J Clin
Virol; 2008, 42: 27-33.
[Abstract]
25. Spinsanti, L, Farías, A, Aguilar, J, Díaz, M P,
Ghisiglieri, S, Bustos, M A, Vilches, N, González, B,
Contigiani M. Risk factors associated with St. Louis
encephalitis seroprevalence in two populations from Córdoba,
Argentina. Trans R Soc Trop Med Hyg; 2007,101:1248-1252.[Abstract]
26. Rotela, C, Spinsanti, L, Lamfri, M, Contigiani, M,
Almiron, W, Scavuzzo, M. Mapping environmental
susceptibility to St.Louis encephalitis virus base don
remote sensing data and geographic information systems.
Vector Borne and Zoonotic Dis (en prensa).
Tablas
Tabla I
Detalle de las características asociadas a las cepas de
virus St. Louis encephalitis aisladas en Argentina.
|
Cepa viral |
Año |
Provincia |
Fuente |
Genotipo1 |
|
Bel |
1963 |
Buenos Aires |
Humanos |
NI |
|
Vil |
1963 |
Buenos Aires |
Humanos |
NI |
|
CorAn 9124 |
1966 |
Córdoba |
Calomys musculinus |
VII |
|
CorAn 9275 |
1966 |
Córdoba |
Mus musculus |
VII |
|
78V-6507 |
1978 |
Santa Fe |
Cx. quinquefasciatus |
VA |
|
79V-2533 |
1979 |
Santa Fe |
Cx. (Cux.)
spp. |
III |
|
AG83-142 |
1982 |
Santa Fe |
Cx. (Cux.)
spp. |
NI |
|
AG84-378 |
1983 |
Santa Fe |
Cx. (Cux.)
spp. |
NI |
|
AG84-A |
1983 |
Santa Fe |
Cx. (Cux.)
spp. |
NI |
|
AG84-B |
1983 |
Santa Fe |
Cx. (Cux.)
spp. |
NI |
|
CbaAr-4005 |
2005 |
Córdoba |
Cx. quinquefasciatus |
III |
|
CbaAr-4006 |
2005 |
Córdoba |
Cx. quinquefasciatus |
III |
1
Clasificación de genotipos según Kramer & Chandler (8)
basado en la secuenciación del gen de la glicoproteína E.
NI: No incluidos en el estudio de genotipificación
Figura 1.
|

