|
TRABAJO ORIGINAL
Resistencia a carbapenemes en
Pseudomonas aeruginosa en un periodo de 10 años en el
Hospital Rawson.
Carbapenems resistance in Pseudomonas aeruginosa during a
10-year period at Rawson Hospital.
Luisa A. Orecchini, Teresa López, Ana Littvik.
Revista Facultad de Ciencias
Medicas 2010; 67(4): 135-140
Unidad de
Microbiología, Hospital Rawson, Córdoba, Argentina.
Bajada Pucara 2025 CP 5014
Introducción
Pseudomonas aeruginosa (Pae) es
un bacilo gram negativo no fermentador que se asocia a
infecciones nosocomiales y en inmunocomprometidos ya que
posee la capacidad de sobrevivir tanto en ambientes húmedos
como antisépticos, aguas de diálisis, etc., como así también
en superficies sólidas, equipos hospitalarios como
ventilación mecánica respiratoria, equipos de diálisis,
etc., [1, 2].
Esto implica un problema
hospitalario ya que la misma puede transmitirse fácilmente a
través de las manos del personal de salud de paciente a
paciente [1, 2].
Por eso es necesario extremar las precauciones para evitar
la transmisión de este microorganismo, ya sea que este
bacilo no fermentador esté colonizando o produciendo
infección.
Pae posee una gran variedad de mecanismos de resistencia
natural, y también una gran capacidad para adquirir
mecanismos nuevos de resistencia o de aumentar la expresión
de su resistencia natural. Este hecho provoca gran
preocupación dentro del ambiente hospitalario, ya que Pae
puede adquirir rápidamente resistencia a muchas drogas
antimicrobianas, y de esta manera perjudicar seriamente la
vida del paciente.
Pae además es capaz de adquirir múltiples mecanismos de
resistencia de manera tal que el perfil de sensibilidad
raramente revela un mecanismo puro de resistencia, sino que
es una sumatoria de varios mecanismos de resistencia.
Resistencia a beta lactámicos
Puede ser por impermeabilidad, bombas de eflujo o
inactivación enzimática.
Por impermeabilidad es resistente natural a macrólidos y
glicopéptidos.
La porina oprF es la principal de Pae, su tamaño es pequeño
por lo tanto muchos antibióticos quedan excluídos, y a su
vez la escasa cantidad de antibiótico que ingresa favorece
el mecanismo de acción de las beta lactamasas que
rápidamente lo hidrolizan
[3].
La porina oprD permite el ingreso de los aminoácidos
básicos, la disminución de su expresión produce resistencia
a imipenem [4, 5, 6],
y sensibilidad disminuida a meropenem
[3, 5, 6].
La pérdida de la porina se debe a la inactivación del gen
que regula su expresión
[5, 6].
La bomba de eflujo MexEF oprN, que confiere resistencia a
quinolonas esta co- regulada con la expresión de oprD, la
expresión de la bomba de eflujo produce una disminución de
la expresión de la porina, por lo tanto también produce
resistencia a imipenem
[3, 5, 6, 7].
En cuanto a las bombas de eflujo MexA MexB oprM es
constitutiva y se expresa en bajo nivel
[3],
confiriendo resistencia a ampicilina, cefalosporinas de
primera y segunda generación. Cuando se aumenta su expresión
confiere resistencia afectando primero a aztreonam y
ticarcilina, luego cefepime y meropenem y luego
piperacilina, ceftazidima doripenem y ciprofloxacina
[4, 5, 6, 8].
La bomba de eflujo Mex CD oprJ produce resistencia a
cefalosporinas de cuarta generación como cefepime
[8].
En relación a la producción de enzimas Pae posee una
cefalosporinasa cromosómica inducible no inhibible por
clavulánico, sulbactam ni tazobactam que le brinda
resistencia natural a ampicilina, amoxicilina clavulánico,
ampicilina sulbactam, cefalosporinas de primera y segunda
generación [3].
Los antibióticos que inducen la
producción de esta enzima son ácido clavulánico y los
carbapenemes, estos últimos son resistentes a la acción de
la enzima [5].
A diferencia de la enterobacterias la frecuencia de mutación
del AmpD es muy baja por lo tanto la derrepresión es
infrecuente, la misma suele verse más frecuentemente en
fibroquísticos, esto podría deberse a la hipermutabilidad
que tienen las cepas de estos pacientes
[6].
Sin embargo es más frecuente su hiperproducción
[5].
Las betalactamasas de espectro ampliado que adquieren mas
frecuentemente son PSE CARB y OXA, otras como TEM 1, TEM 2 y
SHV 1 son poco frecuentes. Brindan resistencia a
piperacilina y se mantienen sensibles a ceftazidima,
aztreonam, y carbapenemes
[5].
Las betalactamasas de espectro extendido encontradas mas
frecuentemente son GES y OXA, y las menos frecuentes son PER
y VEB. Confieren resistencia a penicilinas, cefalosporinas y
monobactames [5].
Las carbapenemasas son betalactamasas que actúan sobre todos
los carbapenemes aparecidos hasta ahora
[8].
Se clasifican en metalo betalactamasas (MBL) si contienen en
su sitio activo una molécula de zinc perteneciendo al grupo
3 de la clasificación de Bush Jacoby y Medeiros; y al grupo
2 serin betalactamasas si contienen un grupo serin en su
sitio activo.
Las MBL son más frecuentes en Pae; son inhibidas por agentes
quelantes de cationes divalentes como EDTA debido a la
presencia de Zn en su molécula. Esta enzima confiere
resistencia todos los betalactámicos excepto a aztreonam
[9].
No es inhibible por ácido clavulánico, sulbactam ni
tazobactam [5, 10].
En 1990 se detectó el primer caso de carbapenemasas (IMP-1)
en Pae en Japón [12]
y luego se describieron
numerosas: VIM, SPM, IMP, GIM
[1, 4, 5, 10, 11]
en diferentes países: Italia (Verona), en Francia, en Brasil
y en Alemania [12].
Objetivos
Determinar los mecanismos de resistencia a carbapenemes en
Pae prevalentes en nuestro medio.
Describir como han evolucionado dichos mecanismos a lo largo
del período considerado.
Materiales y Métodos
Se recolectaron 372 cepas de Pae
de diferentes muestras clínicas durante un período de 10
años comprendido entre 2000 y 2009 De las mismas fueron
seleccionadas 85 cepas que mostraron sensibilidad disminuída
o resistencia a carbapenemes de acuerdo a las normas CLSI
2009 [13],
tomando como punto de corte para sospecha de carbapenemasa
un diámetro de imipenem menor o igual a 21mm.
Las muestras estudiadas fueron analizadas según el protocolo
WHONET (Programa de Vigilancia de resistencia de la OMS)
[14]
y correspondieron a muestras respiratorias, orina, sangre y
catéter.
Fueron identificadas a través de: coloración de Gram,
detección de citocromo oxidasa, demostración de metabolismo
no fermentativo en TSI, utilización de citrato, producción
de arginina dehidrolasa, gelatinasa y pigmento (piocianina).
Se les realizó antibiograma por difusión con discos por el
método de Bauer y Kirby
[15] a los siguientes
antimicrobianos: ceftazidima (30ug); cefepime(30ug);
imipenem(10ug); meropenem (10ug); polimixina (300u);
piperacilina tazobactam (100/10ug); ciprofloxacina (5ug);
piperacilina (100ug); aztreonam (30ug); gentamicina (10ug),
amikacina (30ug). Se interpretaron los resultados de los
halos de inhibición en las categorías de sensible,
intermedio o resistente de acuerdo a las normas CLSI 2009
[13].
Se le colocó un disco de EDTA/SMA, entre los discos de
meropenem e imipenem como screening para detectar la
presencia de MBL.
A las cepas sospechosas de poseer una carbapenemasa se le
realizó el test confirmatorio de resistencia enzimática
Masuda, mediante la técnica descripta por Machiaro y col
[16].
Se hizo una suspensión bacteriana densa en tubos eppendorf
conteniendo 200 uL de buffer fosfato 0.01M PH. A este
inóculo se le agregó 300mg de esferas de zirconio. Se llevó
a vórtex por más de 3 minutos. Luego se centrifugó a más de
6000 rpm por 20 minutos. Se goteó discos de papel de filtro
estériles con el 10uL del sobrenadante. Esto se realizó con
la cepa problema, un control negativo (Pae ATCC 27853) y un
control positivo (Pseudomonas putida productora de MBL
provista por el Servicio de Antimicrobianos del Instituto
Malbrán) En una placa de Mueller Hinton hisopada con un
inóculo 0.5 Mac Farland de Escherichia coli ATCC 25922, se
colocaron los discos de imipenem y ceftazidima. Los discos
con la cepa en estudio, control positivo y negativo se
colocaron a 3 mm por dentro del borde del halo de
sensibilidad registrado en el último control de calidad
interno de la sección antimicrobianos de nuestro hospital.
Resultados
En el período analizado se
aislaron 372 cepas de Pae.
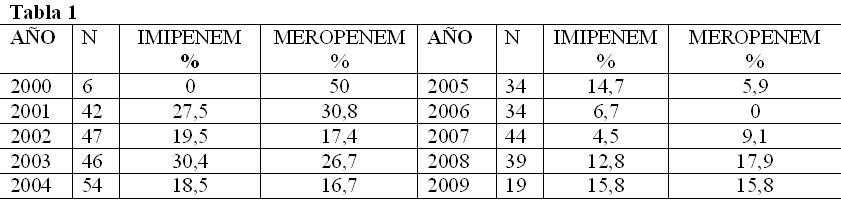
Los perfiles de resistencia para ambos carbapenemes se
observan en la tabla 1.
A todas las cepas con perfil genotípico compatible con
metalo betalactamasa se les realizo la técnica de Masuda
para la detección de resistencia enzimática, y todas fueron
negativas.
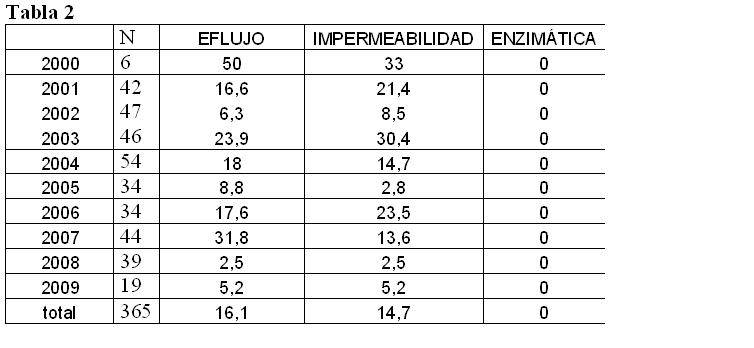
En cuanto a los mecanismos de resistencia a carbapenemes por
eflujo, impermeabilidad y enzimática en el período
considerado se muestran en la tabla 2.
No se detectó resistencia enzimática mediante la técnica de
Masuda.
En los siguientes histogramas se muestran como fueron
variando los porcentajes de resistencia para los
betalactámicos a lo largo del periodo estudiado.
Dichos resultados fueron analizados con el programa WHONET.
El gráfico de resistencia a carbapenemes explica como se
fueron comportando los mecanismos de resistencia a
carbapenemes a lo largo del periodo estudiado.
Discusión
Las betalactamasas de expectro
extendido (TEM, SHV, PER, VEB, y GES) y las metalobeta
lactamasas (incluyendo la familia VIM) se han descripto en
Europa, Asia y America
[17].
La asociación de betalactamasas de espectro extendido y de
metalo betalactamasas se encuentra en raras ocasiones, sin
embargo en Argentina se ha descripto la coexistencia de VIM
y GES [17].
En el hospital Eva Perón Provincia de Buenos Aires,
Argentina, se han detectado en el 2004- 2005 18 cepas con
IMP 13 [18].
En Argentina el número de aislamientos está en ascenso.
Se han publicado un gran número de casos de Pae
multiresistentes; en Texas en 2005-2007 se realizó un
estudio en el cual de 235 aislamiento, 33 fueron cepas
multiresistentes, en cuanto a la resistencia a carbapenemes
, el 44% se debió a hiperexpresion de MexAB, y solo se
encontró un caso de resistencia enzimática por VIM
[19].
En Francia se publicó un trabajo de prevalencia en
Pseudomonas multiresistentes donde se halló que 474
aislamientos, 24 presentaban pérdida de la porina D2 y un
78% de resistencia a meropenem de los cuales un 60%
presentaban hiperexpresión de bombas de eflujo (un 28% del
total de aislamientos)
[20].
En 1990 se detectó el primer caso de carbapenemasas (IMP-1)
en Pae en Japón [12] y luego se describieron numerosas: VIM,
SPM, IMP, GIM [1, 4, 5,
10, 11] en diferentes
países: Italia (Verona), en Francia ,en Brasil y en Alemania
[12].
De esta manera la prevalencia de carbapenemasas se ha
incrementado con el paso del tiempo, diseminándose a Estados
Unidos y Canadá con informes de VIM y IMP
[12].
Se han informado en Australia, y en distintos centros
médicos de la India [12,
21].
En Taiwan el primer aislamiento fue en 2001 de VIM 3. En
esta publicación se encontró una prevalencia de resistencia
enzimática de 1,9% para VIM 2 y de 28,6% para VIM 3, en 308
cepas de Pae resistente a carbapenemes
[22].
Nuestro trabajo demuestra que hay una sumatoria de
resistencias involucrando varios mecanismos. En nuestra
institución a la fecha (2009), la resistencia a carbapenemes
al igual que en la mayor parte del mundo se debe a
mecanismos de impermeabilidad e hiperexpresión de bombas de
eflujo pero a diferencia de otras instituciones de Argentina
y el resto del mundo no hubo resistencia por carbapenemasas.
Conclusiones
En el período estudiado no se
demostró resistencia enzimática a carbapenemes Es probable
que los únicos mecanismos implicados hasta ahora son por
impermeabilidad por pérdida de la porinas y al aumento de
expresión de la bomba de eflujo
La ausencia de resistencia enzimática adquiere gran
relevancia desde el punto de vista epidemiológico ya que la
transmisión horizontal de este mecanismo es muy rápida.
La presencia de carbapenemasas confiere una alta
probabilidad de falla de tratamiento y aumenta la
morbimortalidad. Es importante la detección temprana de las
mismas, dentro de las 72 horas de iniciado el tratamiento.
Es imprescindible contar en las instituciones del cuidado de
la salud, con personal entrenado en la metodología de
detección de estos mecanismos de resistencia y en la
correcta interpretación de una prueba de sensibilidad
antimicrobiana
Bibliografía
[1] Paterson D. The
Epidemiological Profile of Infections with Multidrug-Resistant
Pseudomonas aeruginosa and Acinetobacter Species. Clinical
Infectious Diseases 2006; 43:S43–8.
Full Text
[2] Bertrand X., Bailly P., Blasco G., Balvay P., Boillot
A., Talon D. Large Outbreak in a Surgical Intensive Care
Unit of Colonization or Infection with Pseudomonas
aeruginosa that Overexpressed an Active Efflux Pump.
Clinical Infectious Diseases 2000; 31:e9–14.
Full Text
[3] Livermore D. Multiple Mechanisms of Antimicrobial
Resistance in Pseudomonas aeruginosa: Our Worst Nightmare?.
Clinical Infectious Diseases 2002; 34:634–40.
Full Text
[4] Laupland K., Parkins M., Church D., Gregson D., Louie
T., Conly J., Elsayed S., Pitout J. Population-Based
Epidemiological Study of Infections Caused by Carbapenem-Resistant
Pseudomonas aeruginosa in the Calgary Health Region:
Importance of Metallo-b-Lactamase (MBL)–Producing Strains.
The Journal of Infectious Diseases 2005; 192:1606–12.
Full Text
[5] Bonomo R., Szabo D. Mechanisms of Multidrug Resistance
in Acinetobacter Species and Pseudomonas aeruginosa.
Clinical Infectious Diseases 2006; 43:S49–56.
Full Text
[6] Wolter D., Acquazzino D., Goering R., Sammut P., Khalaf
N., Hanson N. Emergence of Carbapenem Resistance in
Pseudomonas aeruginosa Isolates from a Patient with Cystic
Fibrosis in the Absence of Carbapenem Therapy. Clinical
Infectious Diseases 2008; 46:e137–41.
Full Text
[7] Lister P., Wolter D. Levofloxacin-Imipenem Combination
Prevents the Emergence of Resistance among Clinical Isolates
of Pseudomonas aeruginosa Clinical Infectious Diseases 2005;
40:S105–14.
Full Text
[8] Rice L. Challenges in Identifying New Antimicrobial
Agents Effective for Treating Infections with Acinetobacter
baumannii and Pseudomonas aeruginosa. Clinical Infectious
Diseases 2006; 43:S100–5.
Full Text
[9] Paterson D. Serious Infections in the Intensive Care
Unit: Pseudomonas aeruginosa and Acinetobacter baumannii.
Clinical Infectious Diseases 2006; 43:S41–2.
Full Text
[10] Yoichi Hirakata, Toshiyuki Yamaguchi,a Michiko Nakano,
Koichi Izumikawa, Mariko Mine, Shiho Aoki, Akira Kondoh,
Junichi Matsuda, Mitsukuni Hirayama, Katsunori Yanagihara,
Yoshitsugu Miyazaki, Kazunori Tomono, Yasuaki Yamada,
Shimeru Kamihira, and Shigeru Kohno. Clinical and
Bacteriological Characteristics
of IMP-Type Metallo-b-Lactamase–Producing Pseudomonas
aeruginosa. Clinical Infectious Diseases 2003; 37:26–32.
Full Text
[11] Fritsche T., Sader H., Toleman M., Timothy R. Walsh T.,
Jones R. Emerging Metallo-b-Lactamase–Mediated Resistances:
A Summary Report from the Worldwide SENTRY Antimicrobial
Surveillance Program. Clinical Infectious Diseases 2005;
41:S276–8.
Full Text
[12] Queenan A., Bush K. Carbapenemases: the Versatile
Lactamases. CLINICAL MICROBIOLOGY REVIEWS, July 2007, p.
440–458.
Full text
[13] Clinical and Laboratory Standards Institute.
Performance Standards for Antimicrobial Disk Susceptibility
Testsc Approved Standard. Tenth edition. 2009
Full Text
[14] WHONET. Programa de vigilancia de Resistencia de la
OMS. John Stelling, MD, MPH. Collaborating Centre for
Surveillance of Antimicrobial Resistance Brigham and Women’s
Hospital, Boston.
[15] Bauer, Kirby. Antibiotic susceptibility testing by a
standardized single disk method. Am J Clin Pathol 1966; 45:
493-496.
[16] Marchiaro P., Mussi M., Ballerini V., Pasteran F.,
Viale A., Vila A., Limansky A. Sensitive EDTA-Based
Microbiological Assays for Detection of Metallo-_-Lactamases
in Nonfermentative Gram-Negative Bacteria. JOURNAL OF
CLINICAL MICROBIOLOGY, Nov. 2005, p. 5648–5652.
Full Text
[17] Pasteran F., Faccone D., Petroni A., Rapoport M., Galas
M., Vázquez M., Procopio A. Novel Variant (blaVIM-11) of the
Metallo-_-Lactamase blaVIM Family in a GES-1 Extended-Spectrum-_-Lactamase-Producing
Pseudomonas aeruginosa Clinical Isolate in Argentina.
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Jan. 2005, p.
474–475.
Full Text
[18] Santella G., Cuirolo A., Almuzara M., Palombarani S.,
Sly G., Radice M., Gutkind G. Full Resistance and Decreased
Susceptibility to Carbapenems in IMP-13-Producing
Pseudomonas aeruginosa Isolates from an Outbreak.
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Mar. 2010, p.
1381–1382.
Full Text
[19] Tam V., Chang K., Abdelraouf K., Brioso C., Ameka M.,
McCaskey L., Weston J., Caeiro J., Garey K. Prevalence,
Resistance Mechanisms, and Susceptibility of Multidrug-Resistant
Bloodstream Isolates of Pseudomonas aeruginosa.
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Mar. 2010, p.
1160–1164
Full Text
[20] Rodríguez-Martínez J. M., Poirel L., Nordmann P.
Molecular Epidemiology and Mechanisms of Carbapenem
Resistance in Pseudomonas aeruginosa. ANTIMICROBIAL AGENTS
AND CHEMOTHERAPY, Nov. 2009, p. 4783–4788.
Full Text
[21] Castanheira M., Bell J., Turnidge J., Mathai D., Jones
R. Carbapenem Resistance among Pseudomonas aeruginosa
Strains from India: Evidence for Nationwide Endemicity of
Multiple Metallo-_-Lactamase Clones (VIM-2, -5, -6, and -11
and the Newly Characterized VIM-18). ANTIMICROBIAL AGENTS
AND CHEMOTHERAPY, Mar. 2009, p. 1225–1227.
[22] Sung-Pin Tseng, Jui-Chang Tsai, Lee-Jene Teng and Po-Ren
Hsueh. Dissemination of transposon Tn6001 in carbapenem-non-susceptible
and extensively drug-resistant Pseudomonas aeruginosa in
Taiwan. Journal of Antimicrobial Chemotherapy (2009) 64,
1170–1174.
Full Text

|